This content is also available in:
![]() English
English ![]() Italiano
Italiano ![]() Español
Español ![]() Čeština
Čeština ![]() Magyar
Magyar
Wirusy brodawczaka ludzkiego (Human Papilloma Viruses, HPV) nale?? do rodziny wirusów Papova. Posiadaj? one powinowactwo do komórek nab?onka, indukuj? ich proliferacj?, co prowadzi do rozwoju ?agodnych zmian brodawkowych. Zmiany te stwierdza si? uk?adzie rozrodczym, w górnych drogach oddechowych, przewodzie pokarmowym i na skórze. W oparciu o metody molekularnej hybrydyzacji HPV DNA uzyskanego z k?ykcin i zmian brodawkowych w ró?nych lokalizacjach zidentyfikowano oko?o 70 typów HPV Ka?dy z wirusów wykazuje specyficzno?c, co do miejsca infekcji, a wirusy, które charakteryzuj? si? powinowactwem do tych samych miejsc wydaj? si? by? genetycznie podobne. Stwierdzono, ?e 30 z 70 typów HPV wyst?puje w ?e?skim uk?adzie rozrodczym.
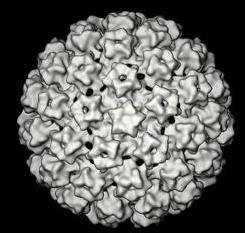
HPV nale?y do wirusów DNA o ?rednicy oko?o 55nm. Genom wirusa zbudowany jest z podwójnej nici DNA, w której wyró?nia si? trzy regiony funkcjonalne: region koduj?cy geny wczesne (E), region koduj?cy geny pózne (L) oraz znajduj?cy si? pomi?dzy nimi d?ugi region kontroluj?cy (Long Control Region, LCR).
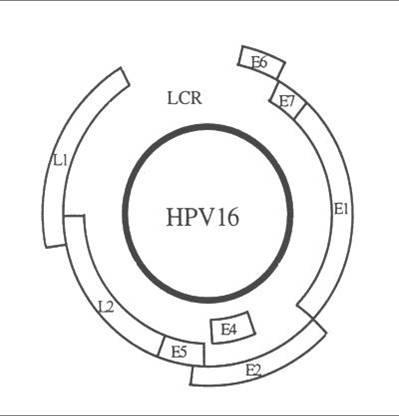
| Gen/region | Funkcja |
|
E1/E2 |
Koduje bia?ka, które kontroluj? funkcj? genów E6 i E7 |
| E4 | Funkcja nieznana, prawdopodobnie kontroluje uwalnianie wirusów z komórki |
| E5 | Koduje bia?ko hydrofobowe, które odgrywa rol? w unie?miertelnieniu komórki |
| E6 | Koduje bia?ka, które hamuj? inhibitory cyklu komórkowego. Produkt genu E6 hamuje bia?ko p53, które promuje apoptoz?. |
| E7 |
Koduje bia?ka, które wi??? si? bia?kiem pRB, co warunkuje progresj? komórki w cyklu komórkowym przy braku sygna?ów mitogennych |
| L1/L2 | Koduje bia?ka strukturalne i odpowiada za tworzenie kompletnych cz?stek wirusa. |
| LCR | Niezb?dny do prawid?owej replikacji wirusa i kontroli ekspresji genów. |
Historia naturalna zaka?enia nab?onka szyjki macicy wirusem HPV i zmiany morfologiczne zwi?zane z tym zaka?eniem.
Wiek w momencie zaka?enia HPV: zaka?enie HPV jest jedn? z najcz??ciej wyst?puj?cych chorób przenoszonych drog? p?ciow?. 75% kobiet przed 50 rokiem ?ycia jest co najmniej raz zaka?onych HPV. Odsetek zaka?e? jest najwy?szy u nastolatek i u kobiet w trzeciej dekadzie ?ycia. Po tym okresie, odsetek zaka?e? zmniejsza si? prawdopodobnie w nast?pstwie odporno?ci przeciwwirusowej. Wirus HPV mo?e przetrwac jako zaka?enie latentne (utajone), które mo?na wykryc tylko metodami diagnostyki molekularnej.
Drogi zaka?enia:: HPV infekuje komórki warstwy podstawnej nab?onka (zwykle w strefie transformacji). Replikacja wirusa wyst?puje w czasie podzia?ów komórek podstawnych. HPV mo?e pozostawac w komórkach warstwy podstawnej w formie latentnej (nieaktywnej) lub ulegac dalszej replikacji w czasie, gdy komórki warstwy podstawnej ulegaj? dojrzewaniu i przemieszczaj? si? w stron? powierzchni nab?onka. Wyk?adnikiem zaka?enia HPV w preparatach histologicznych i cytologicznych jest tzw. efekt cytopatyczny, który charakteryzuje si? koilocytoz?, parakeratoz? i rogowaceniem pojedynczych komórek. Ostre zaka?enie HPV mo?e ulec spontanicznej regresji lub przetrwac, jako zaka?enie utajone, z mo?liwo?ci? ponownej reaktywacji.
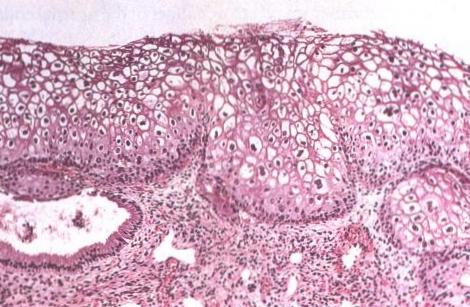
Dowody potwierdzaj?ce koncepcj? roli HPV w rozwoju raka szyjki macicy.
W 1977 roku opublikowano dwie prace, które po raz pierwszy wykaza?y obecno?? HPV w nab?onku szyjki macicy (Meisels & Fortin 1977; Purola & Savia 1977). Autorzy tych prac stwierdzili obecno?? koilocytów w rozmazach z szyjki macicy i w wycinkach z CIN. Opisali oni zmiany w nab?onku szyjki macicy, pochodz?cym ze zmian brodawkowych, które okre?lili, jako “brodawki p?askie” lub “brodawki nie-k?ykcinowe”. Dalsze badania potwierdzi?y obecno?? antygenów wirusa i wirusowego DNA w komórkach nab?onka szyjki macicy i wykaza?y, ?e obecno?? DNA wirusa HPV jest cz?sto zwi?zana ze zmianami o typie CIN1. Ich obserwacje zainicjowa?y dalsze szeroko zakrojone badania nad rol? HPV w karcynogenezie raka szyjki macicy (Coleman and Richman 1983).
Analiza DNA pochodz?cego ze zmian brodawkowych zlokalizowanych w okolicy narz?dów p?ciowych, okolicy odbytu, z nab?onka z cechami CIN i z raka szyjki macicy wykaza?y obecno?? dwóch grup wirusów HPV. Pierwsza grupa to wirusy niskiego ryzyka rozwoju raka szyjki macicy. S? one prawie zawsze zwi?zane z wyst?powaniem CIN1 i k?ykcin zlokalizowanych w okolicy narz?dów p?ciowych i okolicy odbytu. Druga grupa to wirusy wysokiego ryzyka rozwoju raka szyjki macicy. Stwierdza si? je najcz??ciej w nab?onku z cechami CIN2 i CIN3.
Ryzyko rozwoju raka szyjki macicy w zale?no?ci od typu wirusa HPV przedstawiono poni?ej.
Typy wirusa HPV, które wyst?puj? w ?e?skim uk?adzie rozrodczym.
| High risk | 16,18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 68 may lead to Invasive Cancer |
| Low risk | 6, 11, 42, 44, 53, 54, 62, 66 may lead to condylomata |
Potencja? onkogenny wirusów HPV.
Wirusy HPV wysokiego ryzyka charakteryzuj? si? wybitnym potencja?em onkogennym. W badanich in vitro stwierdzono, ?e ludzkie keratynocyty, które przypominaj? komórki docelowe dla HPV, transfekowane wirusami z grupy wysokiego ryzyka (HPV16 i HPV 18) charakteryzuj? si? d?u?szym okresem ?ycia. Keratynocyty transfekowane wirusami niskiego ryzyka (HPV6 i HPV11) dojrzewaj? i umieraj? w tym samym czasie co komórki nie transfekowane. Ponadto ludzkie keratynocyty transfekowane HPV16 i HPV18 wykazuj? cechy morfologiczne podobne do obserwowanych w nab?onku z cechami CIN.
Najbardziej przekonywuj?cym dowodem potwierdzaj?cym rol? HPV w karcynogenezie raka szyjki macicy jest sposób integracji DNA HPV z DNA gospodarza w komórkach nab?onka szyjki macicy. DNA HPV w warunkach prawid?owych, w k?ykciach i w nab?onku z cechami CIN1 wyst?puje w postaci episomalnej (tj. bez zwi?zku z genomem gospodarza). DNA HPV w komórkach raka szyjki macicy jest zintegrowany z DNA gospodarza. Integracja z HPV DNA z DNA gospodarza powoduje zaburzenie ekspresji genów E1 i E2, które w warunkach prawid?owych kontroluj? geny E6 i E7 oraz zaburzenie otwartych ramek odczytu genów E6 i E7. Bia?kowe produkty genów E6 i E7 hamuj? funkcj? genów supresorowych (TP53 i RB), co prowadzi do omini?cia punktów kontroli cyklu komórkowego (G1 i G2) i niekontrolowanej proliferacji.
Integracj? HPV DNA z DNA gospodarza stwierdza sie we wszystkich rakach zawieraj?cych HPV16 i HPV18. Stanowi to najwa?niejszy dowód na znaczenie HPV16 i HPV18 w patogenezie raka szyjki macicy.
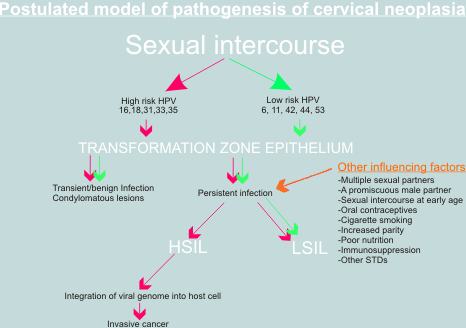
Znaczenie innych czynników.
Wiadomym jest, ?e zaka?enie HPV wysokiego ryzyka lub nawet integracja HPV16 z DNA gospodarza nie jest wystarczaj?ca do pe?nej transformacji z?o?liwej komórek nab?onka szyjki macicy. Zaka?enie uk?adu rozrodczego HPV16 jest stosunkowo cz?ste, a rak szyjki macicy rzadki. Ponadto integracj? HPV DNA stwierdza si? tak?e w niektórych przypadkach k?ykcin i w nab?onku z cechami CIN. W patogenezie raka szyjki macicy mog? odgrywac rol? tak?e inne czynniki, takie jak: upo?ledzona odpowiedz immunologiczna, przetrwa?e zaka?enie wirusowe, palenie papierosów, przyjmowanie doustnych ?rodków antykoncepcyjnych. Wy??czenie genów supresorowych (delecja, mutacja) lub aktywacja onkogenów mo?e tak?e odgrywac rol? w patogenezie raka szyjki macicy. W badaniach na liniach komórek raka szyjki macicy in vitro stwierdzono mutacje RAS, FOS i innych onkogenów, ale znaczenie tych mutacji in vivo nie jest jeszcze w pe?ni poznane. Badania nad rol? HPV w patogenezie raka szyjki macicy stanowi? teoretyczn? przes?ank? do wprowadzenia metod zapobiegania rakowi szyjki macicy (szczepienia), nowych metod wczesnego wykrywania (skryning HPV DNA) i nowych sposobów leczenia (leki przeciwwirusowe).
Zwi?zek mi?dzy HPV a rakiem szyjki macicy.. Ryzyko wzgl?dne jest bardzo wysokie i porównywalne z ryzykiem wyst?pienia pierwotnego raka w?troby w przebiegu przewlek?ego wirusowego zapalenia w?troby typu B. Jest ono tak?e wy?sze ni? w przypadku palenia papierosów i ryzyka rozwoju raka p?uca.